
Modifications phosphonates inédites dans la bave du vers de veloursUnprecedented phosphonate modifications in velvet worm slime
Co-premier auteur (expériences principales). Grâce à une sonde MAS-DNP triple résonance ¹H-¹³C-³¹P unique en Amérique du Nord, découverte de liaisons carbone-phosphore atypiques (phosphonates) décorant les glycoprotéines de la bave du vers de velours, une classe de biomolécule jamais décrite. Repris par Québec Science, ACS News et Radio-Canada (« Découverte », « Les Années Lumière »).Co-first author (led most experiments). Using a unique North-American ¹H-¹³C-³¹P triple-resonance MAS-DNP probe, discovered atypical carbon-phosphorus bonds (phosphonates) decorating glycoproteins in velvet worm slime, a never-before-reported biomolecule class. Covered by Québec Science, ACS News and Radio-Canada (Découverte, Les Années Lumière).
Les vers de velours (Onychophora) forment un groupe ancien d'invertébrés terrestres, dont les deux grandes lignées se sont séparées il y a environ 380 millions d'années. Pour capturer leurs proies, ils projettent une bave collante depuis des papilles situées de chaque côté de leur tête. Ce gel, principalement fait de protéines, se transforme en fibres solides sous l'effet du cisaillement et du séchage rapide ; les efforts de la proie pour se débattre accélèrent même ce durcissement, jusqu'à obtenir des fibres aussi rigides que du Nylon.Velvet worms (Onychophora) are an ancient group of terrestrial invertebrates, whose two major lineages diverged roughly 380 million years ago. To capture prey, they project sticky slime from papillae on either side of their head. This protein-rich gel turns into solid fibres under mechanical shearing and rapid drying; the prey's own struggling even speeds up this hardening, producing fibres as stiff as Nylon.
Epiperipatus barbadensis éjectant sa bave pour capturer une proie ; la bave durcit en quelques secondes au contact de l'air.Epiperipatus barbadensis ejecting slime to capture prey; the slime hardens within seconds on contact with air.
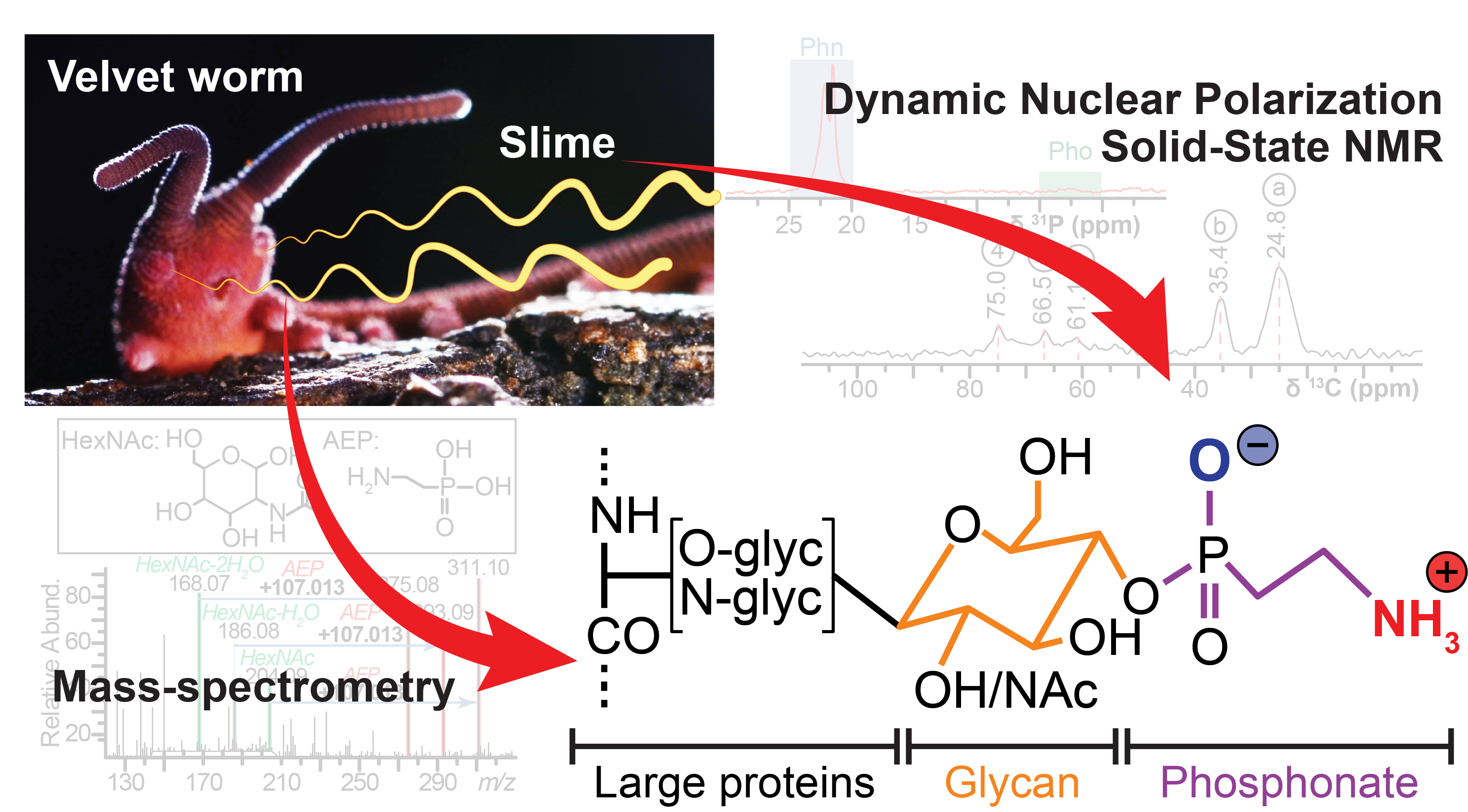
La bave du vers de velours n'est pas isotopiquement marquée, ce qui rend ses signaux RMN extrêmement faibles. Décrire sa composition moléculaire exacte nécessitait une sensibilité hors d'atteinte des approches RMN classiques.Velvet worm slime is not isotopically labelled, making its NMR signals extremely weak. Describing its exact molecular composition required sensitivity beyond standard NMR approaches.
Développement et utilisation d'une sonde MAS-DNP triple résonance ¹H-¹³C-³¹P, unique en Amérique du Nord, combinée à de la spectrométrie de masse, pour sonder des signaux de très faible intensité dans des échantillons non marqués.Development and use of a ¹H-¹³C-³¹P triple-resonance MAS-DNP probe, unique in North America, combined with mass spectrometry, to probe very low-intensity signals in unlabelled samples.
Découverte de liaisons carbone-phosphore (phosphonates) inédites décorant les glycoprotéines de la bave, la première description de ce type de modification dans une sécrétion (et non une paroi) d'animal terrestre, une preuve de la parenté évolutive de deux lignées de vers de velours séparées depuis 380 millions d'années.Discovery of unprecedented carbon-phosphorus bonds (phosphonates) decorating slime glycoproteins, the first description of this modification type in a terrestrial animal's secretion (not a cell wall), and evidence of evolutionary kinship between two velvet worm lineages separated for 380 million years.
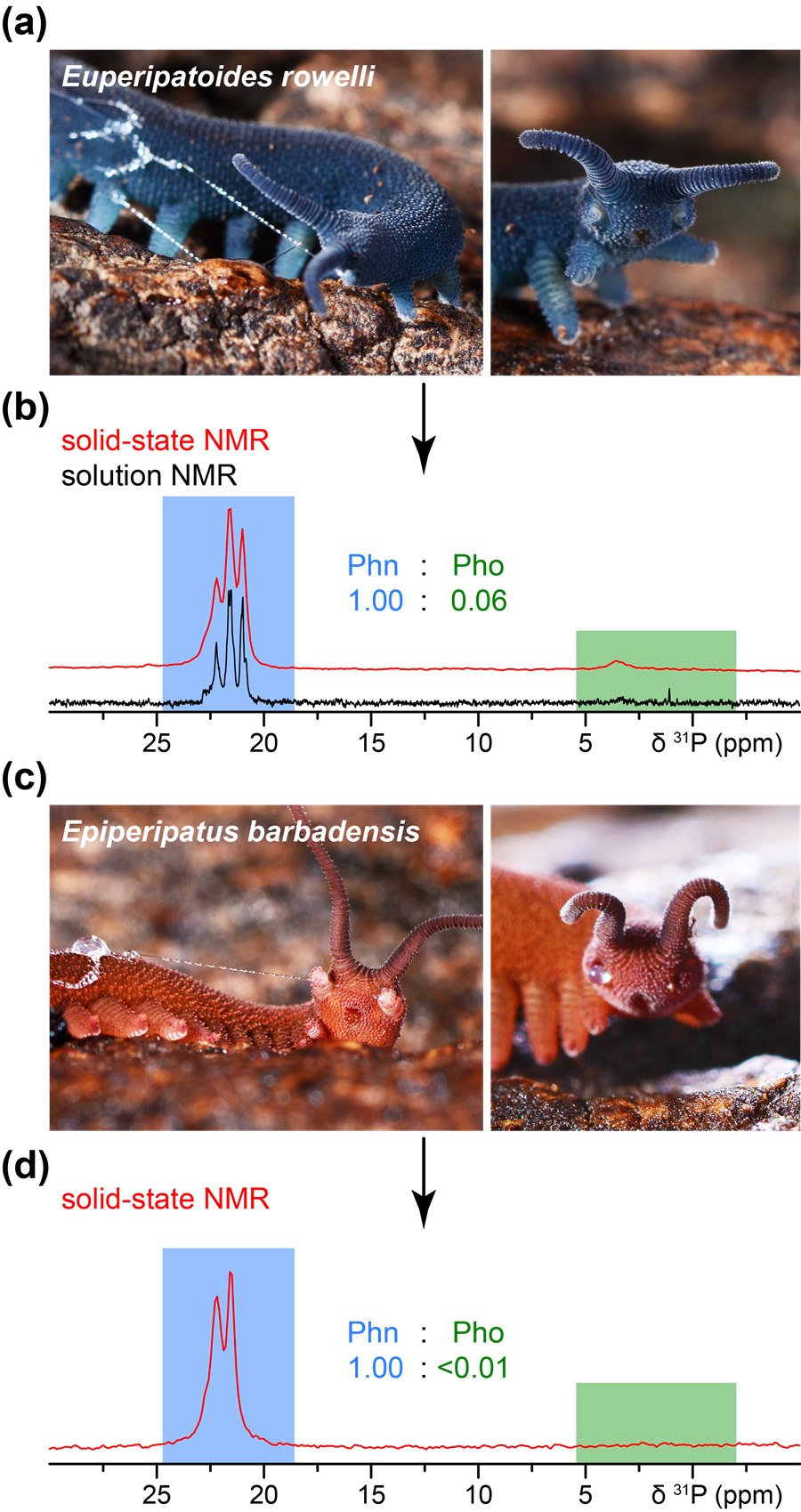
(a) Euperipatoides rowelli et (c) Epiperipatus barbadensis. (b, d) Spectres RMN ³¹P en solution (noir) et RMN de l'état solide (rouge) : chez E. rowelli, le ratio phosphonates (Phn, zone bleue) / phosphates (Pho, zone verte) est de 1,00:0,06, soit environ 17 fois plus de phosphonates que de phosphates ; chez Ep. barbadensis, les phosphates sont quasiment absents (ratio < 0,01), ne laissant que le signal des phosphonates.(a) Euperipatoides rowelli and (c) Epiperipatus barbadensis. (b, d) Solution (black) and solid-state (red) ³¹P NMR spectra: in E. rowelli, the phosphonate (Phn, blue region) to phosphate (Pho, green region) ratio is 1.00:0.06, about 17 times more phosphonate than phosphate; in Ep. barbadensis, phosphates are essentially absent (ratio <0.01), leaving only the phosphonate signal.
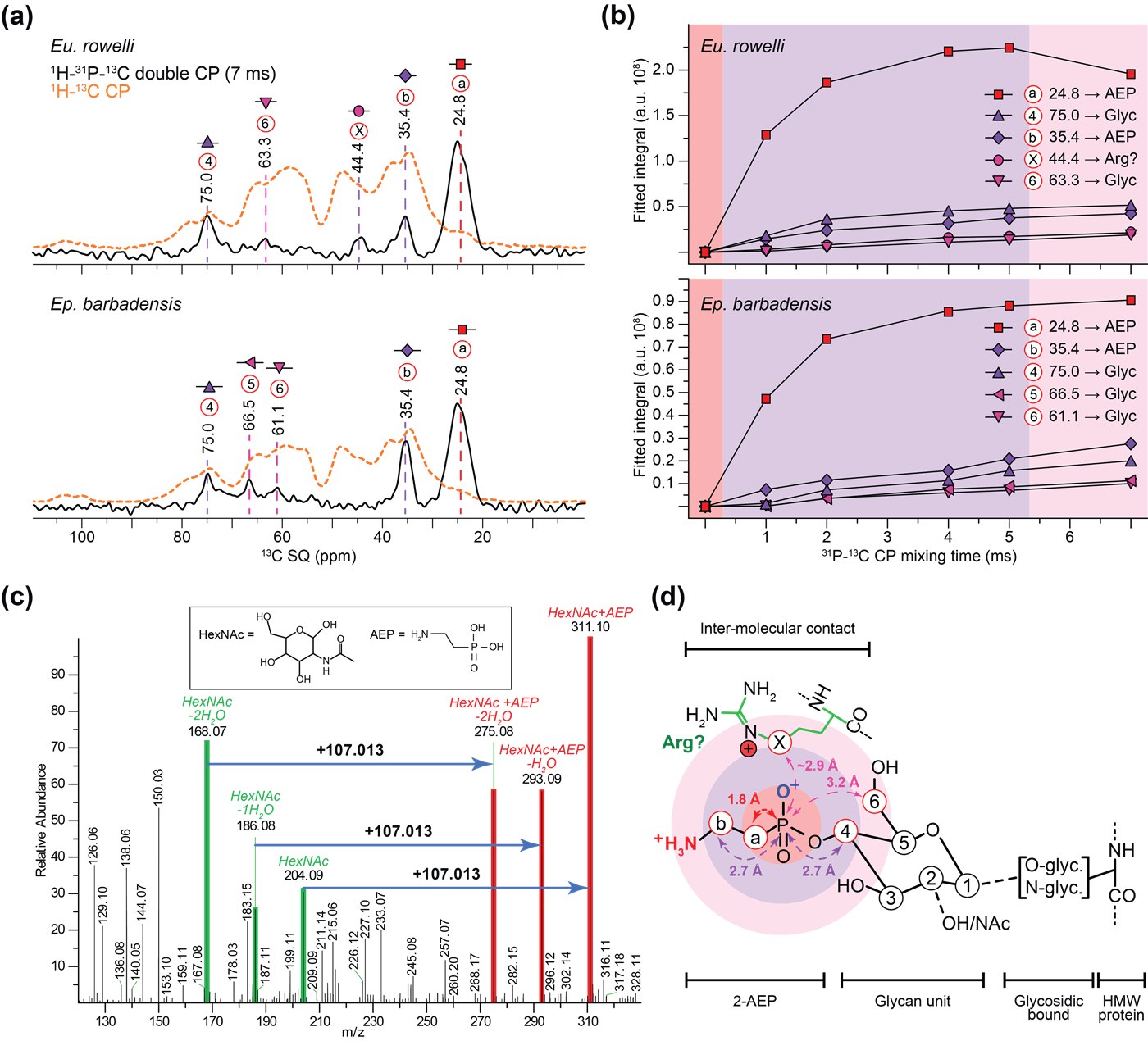
(a) Spectres MAS-DNP ¹H-³¹P-¹³C (double CP, noir) comparés au spectre direct ¹H-¹³C CP (orange) : seuls les carbones proches du ³¹P apparaissent dans le premier, révélant leur proximité avec le phosphonate. (b) Intensité de ces mêmes pics selon le temps de contact CP : les carbones les plus proches apparaissent en premier, les plus éloignés ensuite, ce qui permet de cartographier les distances. (c) Spectrométrie de masse en tandem (HCD-MS/MS) de peptides trypsiques : des fragments HexNAc portant une masse additionnelle de 107,013 confirment la modification par le 2-AEP (2-aminoéthylphosphonate). (d) Schéma du glycane phosphonaté greffé sur une protéine de haut poids moléculaire de la bave, avec les distances estimées par MAS-DNP et calculs DFT.(a) MAS-DNP ¹H-³¹P-¹³C double-CP spectra (black) compared to the direct ¹H-¹³C CP spectrum (orange): only carbons close to ³¹P appear in the former, revealing their proximity to the phosphonate. (b) Intensity of these same peaks as a function of CP contact time: the closest carbons build up first, more distant ones later, allowing distances to be mapped. (c) Tandem mass spectrometry (HCD-MS/MS) of tryptic peptides: HexNAc fragments carrying an extra 107.013 mass unit confirm the 2-AEP (2-aminoethylphosphonate) modification. (d) Schematic of the phosphonated glycan grafted onto a high-molecular-weight slime protein, with distances estimated by MAS-DNP and DFT calculations.